
This is an essay which I submitted as part of a university physiology module, it’s a lot drier than my normal style of writing but hopefully still readable and useful.
Adaptations to the triceps surae muscle group in response to plyometric training.
The stretch shortening cycle (SSC) has been described as a musculotendinous unit (MTU) being pre-stretched before a concentric action, the elastic effect of which acts to improve movement efficiency and performance compared to purely concentric muscle actions, (Fouré, Nordez, McNair, & Cornu, 2011; Turner & Jeffreys, 2010). Plyometric exercises closely resemble the ballistic movements patterns found in athletic performances such as jumping (Vissing et al., 2008)and are theorised to enhance an athlete's utilisation of the SCC (Turner & Jeffreys, 2010), therefore many coaches incorporate plyometric exercises into their athletes' training in order to improve performance. Clark, Ryan and Weyand (2017) were able to predict vertical ground reaction forces during running by creating a two-mass model, separating the lower limb below the knee from the rest of the body mass. Their model suggests that the impulse experienced by the lower limb from contact with the ground is different to the vertical accelerations of the rest of the body mass during the ground contact period. How the lower limbs deal with the forces experienced, known as the force-deformation characteristics, influences many athletic tasks such as running, jumping or changing direction (Brazier, Maloney, Bishop, Read, & Turner, 2017). Furthermore during jumping tasks coaches have often placed a high level of importance on the Achilles tendon (Earp et al., 2011) due to the unique elastic energy storage and transmission characteristics of musculotendinous units with long tendons during the SSC (Hirayama et al., 2017). This essay will therefore explore how the musculotendinous units of the triceps surae muscle group adapt in response to plyometric training.
Mechanotransduction is the process by which the body converts mechanical loading into a cellular response (Khan & Scott, 2009). This response creates an adaptation, or structural change, in order for the body to be able to cope with the demands being placed upon it. This means that the Achilles tendon, for example, will respond differently depending to the loading duration, magnitude and type that it experiences. Coaches can use this specific adaptation to imposed demands (the SAID principle) to design training drills to optimise the adaptive response to the needs of the athlete and their sport (Mattacola & Dwyer, 2002). Different plyometric exercises will impose different demands and therefore elicit different responses, depending on the design of the exercise. For example, plyometric exercises are often categorised as being fast or slow depending on whether the ground contact time is faster or slower than 250ms (Jeffreys & Turner, 2010) and Bosco, Tihanyi, Komi, Fekete and Apor (1982) argue that athletes with a high proportion of fast twitch fibres will benefit the most from jumping strategies that utilise high speed, low amplitude movements, whilst athletes with proportionally more slow twitch fibres are able to maintain cross-bridge attachments longer and may potentially have a better utilisation of elastic energy during slower plyometric activities. Similarly Earp et al. (2011) reported peak vertical ground reaction forces of 2678 ± 657N for a drop jump (DJ) which were significantly higher (p<0.05) than the 2250 ± 413N recorded for a countermovement jump (CMJ). This difference in loading will, according to the theories of mechanotransduction and the SAID principle, elicit different responses which should be considered when looking at research into the adaptations caused by plyometric training. Kubo, Ishigaki and Ikebukuro (2017) found no change in the Achilles stiffness in 11 sedentary males after twelve weeks of plyometric training, however the subjects performed the exercises on a sledge apparatus. Gruber, Kramer, Mulder and Rittweger (2019) also investigated plyometrics using a sled apparatus and again stated that the tissue adaptation is driven by the magnitude of tissue strain. In their case they were looking at ways of minimising bone and muscle mass loss in astronauts during space flight and proposed plyometrics as a method for generating high forces and therefore large training effects. The design of their sledge used pressurised tubes creating a counterforce which was greater than the forces normally experienced in microgravity, it is not clear if the forces experienced on the sledge used in the Kubo et al. (2017) study where significantly greater than the gravitation forces to which the subjects were habituated, which may be the reason for their not finding a training effect in the tendon.
Whilst they didn't find any change in tendon stiffness the Kubo et al. (2017) study did find a significant increase in the thickness of the triceps surae muscles following the plyometric training (5.7 ± 2.6%, p<0.001). The measurement was reported however, as a mean value for medial gastrocnemius, lateral gastrocnemius and soleus thicknesses combined, which Fouré, Nordez, Guette and Cornu (2009) argue against as they suggest that the different anatomical attachment sites, bi-articular or mono-articular structures and differing roles of the different muscles mean that they should respond differently to the training protocol. The hypothesis put forward by Fouré et al. (2009) that the soleus has a mainly postural role and therefore contains more slow titch fibres which respond differently to the fast twitch, movement orientated fibres found in the gastrocnemius is in agreement with the findings of Bosco et al. (1982) regarding the different muscle types being best suited to different plyometric exercises, as discussed previously. Vissing et al. (2008) also found evidence of muscular hypertrophy following a plyometric training intervention on untrained subjects, but not to the same degree as in conventional resistance training. In contrast Kyröläinen et al. (2005) conducted a 15 week study with a mixture of plyometric and ballistic exercises and found no significant changes in muscle fibre type, fibre size or myosin heavy chain isoform composition. Markovic and Mikulic (2010) performed a meta-analysis of adaptations to plyometric training and concluded that whilst there was evidence of hypertrophy in type I, IIa and IIx fibres following plyometric training the results were modest and lower than would be found in traditional weight training, furthermore the evidence for whether changes in muscle fibre type could be induced by plyometric training were inconclusive.
In addition to hypertrophy and fibre type changes the length and pennation angles of the muscle fibres can also be a factor in muscular performance. Earp et al. (2011) found in their cross-sectional study that the subjects who developed the greatest ground reaction forces during a DJ landing had the greatest pennation angles in the gastrocnemius. They state that a greater pennation angle and shorter fascicle lengths are better able to tolerate the large eccentric loads generated in a DJ as the forces are dissipated by a factor of cosine of the angle of pennation, meaning the strain forces are distributed to a greater surface area along the aponeurosis. Conversely a shallower pennation angle and longer fascicle lengths are theorised to allow for faster contraction velocities and the subjects with these characteristics produced a greater rate of force development (RFD) in the early stages of a countermovement jump (CMJ). The explanation that they put forward for this is that during the countermovement the knee is rapidly flexed, which would reduce loading on the gastrocnemius, however the gastrocnemius has to increase the tension in the Achilles tendon to ensure an efficient SSC for the jump and so to keep up with the rapid length changes it needs a high contraction velocity. As the soleus does not cross the knee it does not have to deal with this tension loss during knee flexion and so this again supports the theories of Bosco et al. (1982) and Fouré et al. (2009) that the gastrocnemius and soleus muscles should be considered separately when looking at the adaptations to plyometrics.
Earp et al. (2011) concluded that there is an optimal muscle architecture, depending on the eccentric load and phase of the jump. As these characteristics are trainable we can assume, given the theories of mechanotransduction and the SAID principle, that the muscle will adapt to optimise it's architecture in response to the training loads it experiences. Earp et al. (2011) however state that only heavy strength training has actually been shown to increase pennation angle and therefore coaches looking to create this adaptation in order to improve the eccentric forces that an athlete can withstand (for example being able to tolerate greater drop heights in the DJ) should use this in the athlete's training program. Turner and Jeffreys (2010) also state that strength training should precede plyometric training to ensure sufficient strength is present for the task, but that the muscle characteristics are optimised for sports performance by plyometric training.
As previously mentioned stiffness of the lower limb is thought to be key determinant of athletic performance as it affects the efficient transfer to the ground of forces generated higher up the kinetic chain and efficient usage of the SSC, both of which are thought to be optimised by plyometric training (Fouré et al., 2009; Fouré, Nordez, & Cornu, 2010; Jeffreys & Turner, 2010). Optimised is the key word here, as Brazier et al. (2017) state that there is an optimal level of stiffness - too high and the peak forces and loading rates could lead to an increase in bony injuries such as stress fractures, too low and there is an increased risk of soft-tissue injuries. They state that what is deemed optimal depends on the demands of the sport and the physical profile of the athlete (body mass, limb length etc.), once again in agreement with the principles of specific adaptation.
Stiffness is not consistent across the entire MTU which led Hill (1938) to create a three component model to describe it, consisting of the active muscle discussed above, termed the contractile element (CE), a parallel elastic component (PEC) containing the sarcolemma and muscle fascia and finally a series elastic component (SEC) consisting of the tendons.

Figure 1. Hill (1938) 3 component MTU model. CE = contractile element, PEC = parallel elastic component, SEC = series elastic component.
The model predicts that as a muscle fibre contracts the force will first be transmitted to the SEC which acts as a spring. When enough force is generated the slack will be taken out of the spring and the forces transmitted externally (Wilson & Flanagan, 2008), the difference in time between the activation of the muscle and the generation of external force being termed the electromechanical delay (EMD). During the rapid movements found in many sport skills (such as jumping or plyometric training) this relationship between the CE and SEC allows the muscle to contract with an optimised force-velocity relationship whilst the elastic properties of the SEC can react to and express forces quickly (Kubo, Morimoto, Komuro, Yata, et al., 2007). In addition the elastic properties of the SEC are thought to improve locomotion efficiency by reducing the metabolic cost of motion (Turner & Jeffreys, 2010). The triceps surae have a relatively unique arrangement amongst the muscles used in locomotion in that the SEC (the Achilles tendon) is very long compared to the CE (the gastrocnemius and soleus muscles) in order to maximise the usage of elastic energy (Wilson & Flanagan, 2008), as illustrated in figure 2.
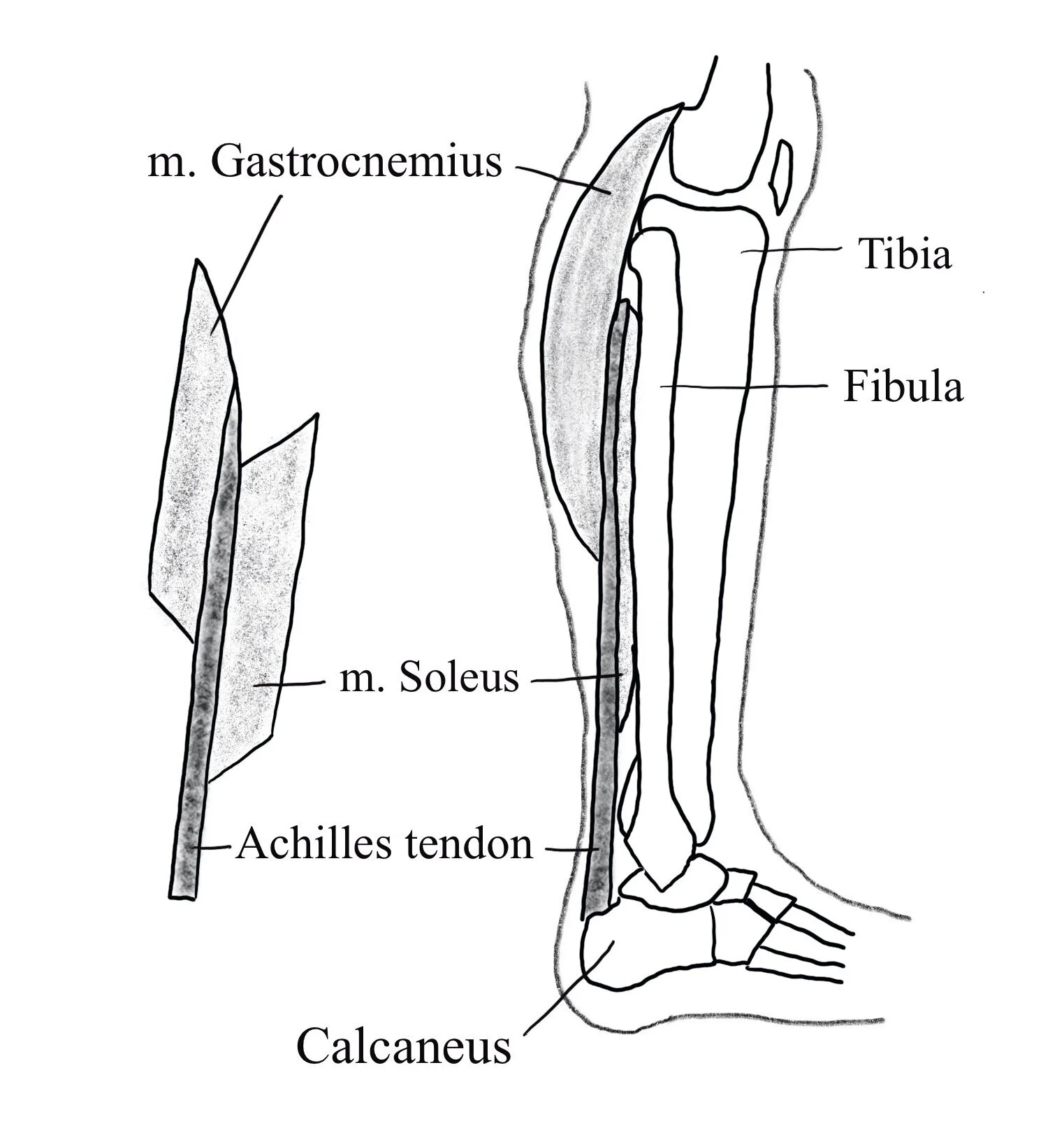
Figure 2. Lateral view of the triceps surae. Adapted from Shan et al. (2019).
Fouré et al. (2010) and Fouré et al. (2011) ran studies looking at the changes in Achilles tendon stiffness following 14 weeks of plyometric training in active sports people and found that the stiffness increased. In contrast Fouré et al. (2009)used subjects from explosive sports (such as volleyball) and found no change, suggesting that the explosive athletes were already well adapted to the demands. Unfortunately there were differences in the training protocols used in the different studies which may have affected this conclusion. The Fouré et al. (2010) and Fouré et al. (2011) studies used a 14 week intervention with a total of 6800 jumps whereas the Fouré et al. (2009) study used an 8 week intervention with a total of 3200 jumps.
Kubo et al. (2017) found that the extensibility of the Achilles tendon was velocity dependant and that plyometric training optimised this for ballistic movements and the SSC. This may also explain why some authors do not find a change in tendon properties following plyometric training, by using the common ramp testing protocol for example, they may miss the changes seen at higher velocities.
Another reason why researchers may miss the expected changes in tendon properties is that the training interventions tend to be relatively short. In a meta-analysis Bohm et al. (2015) reported weighted average effect sizes of 0.91 for interventions over 12 week and 0.81 for shorter durations. Kubo, Ikebukuro, Yata, Tsunoda and Kanehisa (2010) agree, though their research was looking at single joint isometric training rather than plyometrics. They tested tendon stiffness regularly throughout the training intervention and found that the stiffness increases only reached statistical significance at the end of the three month intervention.
In summary, evidence for changes in the stiffness of the Achilles tendon following plyometric training remain inconclusive (Fouré et al., 2010; Markovic & Mikulic, 2010; Turner & Jeffreys, 2010), a number of authors have found increases (Fouré et al., 2011; Wu et al., 2010) whilst others have found no change (Kubo et al., 2017). Most authors are in agreement though, that any changes that are seen following plyometric training are not due to a change in the cross sectional area of the tendon, but rather it's structural composition (Bohm, Mersmann, & Arampatzis, 2015; Fouré et al., 2010, 2011; Kubo, Morimoto, Komuro, Yata, et al., 2007), though again the exact nature of this change (alterations to collagen cross-bridges, hydration status etc.) is unclear.
The final component of Hill's model is the PEC, the stiffness of which is 100 times less than that of the SEC and so during a passive stretch it is the PEC which deforms and stores elastic energy. During a contraction however the stiffness of the CE and surrounding PEC exceeds that of the SEC and therefore it is the tendons which deform and become the main site for elastic energy storage (Turner & Jeffreys, 2010; Wilson & Flanagan, 2008). It is the control of this interplay, along with that of the agonist/antagonist muscle actions by the central nervous system (CNS) which regulates the overall limb/joint stiffness that is so important to efficient use of the SSC and to overall athletic performance (Chmielewski, Myer, Kauffman, & Tillman, 2006).
Davies, Riemann, & Manske (2015) state that plyometric training enhances neuromuscular coordination which results in the performance increases seen. One part of this is an increased excitability of the muscle spindles which in response increases the activation of the muscle as a protective reflex action (Markovic & Mikulic, 2010). At the same time Golgi Tendon Organs (GTO) which detect stretch in the SEC and act to decrease muscle activation, again as a protective mechanism, are desensitised. This coupled excitation of the muscle stretch reflex and desensitisation of the GTOs together allow for a more forceful contraction. Wu et al. (2010) found significant increases in EMG activity (p<0.001) in the soleus following eight weeks of plyometric training, but not in the gastrocnemius. The authors state that as the soleus is mono-articular it experiences a greater stretch during a countermovement than the bi-articular gastrocnemius and therefore a greater stretch reflex. This adds further evidence to the differences between the soleus and gastrocnemius muscles previously discussed.
Wu et al. (2010) were also one of the research groups who found an increase in tendon stiffness following plyometric training. This, together with the increased activation levels combined to form a decrease in EMD, in response to which they postulate that the body may reorganise movement patterns. Once again changes in EMD are not a universal finding, but supporting evidence for coordination changes comes from Hirayama et al. (2017) who were looking at neuro-muscular behaviour during depth jumping. They found that after a plyometric training program the EMG values for the gastrocnemius increased during the breaking phases of a jump and decreased during the early acceleration phase. During the late breaking and early acceleration phases the muscle fibres were isometric, maximising the usage of elastic energy and optimising the force-velocity relationship of the muscle fibres, as discussed previously. During the early breaking phase the fascicles were shortening after the training intervention whereas prior to it the fascicles lengthened throughout the whole of the breaking phase. In addition to these changes the activity of the antagonist muscle (the tibialis anterior) was seen to decrease during the early breaking, late breaking and late acceleration phases. Increased usage of elastic energy and decreased antagonist action led to increased jump performance (impulse increased from 168 ± 21N.s to 192 ±20 N.s, p<0.001, contact time decreased from 0.365 ± 0.068s to 0.310 ±00.43s, p=0.01). Kubo, Morimoto, Komuro, Tsunoda, et al. (2007) also found similar patterns when looking at the differences between CMJs, DJs and squat jumps (SJ) - as the loading forces experienced during the jumps increased the EMG values during the breaking phases also increased. During the concentric phases however, there were no differences between DJ and SJ EMG values but CMJ was significantly higher (p<0.001). Their conclusion is that stretch reflexes increase muscle activation (firing rate and number of motor units) to create a stronger concentric contraction in the CMJ whereas the higher loading in the DJ is handled by keeping the muscle fascicles in the more advantageous isometric range and increasing the usage of elastic energy from the SEC. Davies et al. (2015) state that these neuro-muscular control adaptations account for the majority of the performance increases seen during the first 6 to 8 weeks of plyometric training, with morphological adaptations occurring more slowly.
In conclusion, plyometric training has been shown to be an effective training methodology for optimising usage of the SSC for athletic performance. A lot of focus within plyometric training has been placed on the triceps surae muscle group due to their role in locomotion and their anatomy, with a long Achilles tendon being optimal for the utilisation of elastic energy. Mechanotransduction and the SAID principles say that the triceps surae should adapt to the loads placed upon them by plyometric training, however in the first 6 to 8 weeks the observed performance increases come mainly from improvements to neurological control mechanisms. Morphological adaptations such as hypertrophy or changes to tendon stiffness happen over a much longer time frame and not to the same extent as seen in other training methodologies. Therefore a coach looking for a specific adaptation, an increase to Achilles tendon stiffness following a tendinopathy for example, would be better served with other training methodologies (in this case heavy resistance training). Some authors also argue that resistance training should precede plyometric training to create a base of strength to help the body cope with the high forces experienced in plyometric training, but there is an agreement that in order to optimise the triceps surae's use of the SSC for athletic actions such as running and jumping plyometric training should form part of an athlete's training plan.
References.
Bohm, S., Mersmann, F., & Arampatzis, A. (2015). Human tendon adaptation in response to mechanical loading: a systematic review and meta-analysis of exercise intervention studies on healthy adults. Sports Medicine - Open, 1(1).
Bosco, C., Tihanyi, J., Komi, P. V., Fekete, G., & Apor, P. (1982). Store and recoil of elastic energy in slow and fast types of human skeletal muscles. Acta Physiologica Scandinavica, 116, 343–349.
Brazier, J., Maloney, S., Bishop, C., Read, P., & Turner, A. (2017). Lower Extremity Stiffness: Considerations for Testing, Performance Enhancement and Injury Risk. Journal of Strength and Conditioning Research, 33(4), 1156–1166.
Chmielewski, T. L., Myer, G. D., Kauffman, D., & Tillman, S. M. (2006). Plyometric exercise in the rehabilitation of athletes: Physiological responses and clinical application. Journal of Orthopaedic and Sports Physical Therapy, 36(5), 308–319.
Clark, K. P., Ryan, L. J., & Weyand, P. G. (2017). A general relationship links gait mechanics and running ground reaction forces. Journal of Experimental Biology, 220(2), 247–258.
Davies, G., Riemann, B. L., & Manske, R. (2015). Current Concepts of Plyometric Exercise. International Journal of Sports Physical Therapy, 10(6), 760–786.
Earp, J. E., Kraemer, W. J., Prue, C., Joseph, M., Volek, J. S., Maresh, C. M., & Newton, R. U. (2011). Influence of muscle-tendon unit structure on rate of force development during the squat, countermovement, and drop jumps. Journal of Strength and Conditioning Research, 25(2), 340–347.
Fouré, A., Nordez, A., Guette, M., & Cornu, C. (2009). Effects of plyometric training on passive stiffness of gastrocnemii and the musculo-articular complex of the ankle joint. Scandinavian Journal of Medicine and Science in Sports, 19(6), 811–818.
Fouré, Alexandre, Nordez, A., & Cornu, C. (2010). Plyometric training effects on Achilles tendon stiffness and dissipative properties. Journal of Applied Physiology, 109(3), 849–854.
Fouré, Alexandre, Nordez, A., McNair, P., & Cornu, C. (2011). Effects of plyometric training on both active and passive parts of the plantarflexors series elastic component stiffness of muscle-tendon complex. European Journal of Applied Physiology, 111(3), 539–548.
Gruber, M., Kramer, A., Mulder, E., & Rittweger, J. (2019). The importance of impact loading and the stretch shortening cycle for spaceflight countermeasures. Frontiers in Physiology, 10(MAR).
Hill, A. (1938). The heat of shortening and the dynamic constants of muscle. Proceedings of the Royal Society of London. Series B - Biological Sciences, 126(843), 136–195.
Hirayama, K., Iwanuma, S., Ikeda, N., Yoshikawa, A., Ema, R., & Kawakami, Y. (2017). Plyometric training favors optimizing muscle-tendon behavior during depth jumping. Frontiers in Physiology, 8(JAN), 1–9.
Khan, K. M., & Scott, A. (2009). Mechanotherapy: how physical therapists’ prescription of exercise promotes tissue repair. British Journal of Sports Medicine, 43(4), 247–252.
Kubo, K., Ikebukuro, T., Yata, H., Tsunoda, N., & Kanehisa, H. (2010). Time course of changes in muscle and tendon properties during strength training and detraining. Journal of Strength and Conditioning Research, 24(2), 322–331.
Kubo, K., Ishigaki, T., & Ikebukuro, T. (2017). Effects of plyometric and isometric training on muscle and tendon stiffness in vivo. Physiological Reports, 5(15), 1–13.
Kubo, K., Morimoto, M., Komuro, T., Tsunoda, N., Kanehisa, H., & Fukunaga, T. (2007). Influences of tendon stiffness, joint stiffness, and electromyographic activity on jump performances using single joint. European Journal of Applied Physiology, 99, 235–243.
Kubo, K., Morimoto, M., Komuro, T., Yata, H., Tsunoda, N., Kanehisa, H., & Fukunaga, T. (2007). Effects of plyometric and weight training on muscle-tendon complex and jump performance. Medicine and Science in Sports and Exercise, 39(10), 1801–1810.
Kyröläinen, H., Avela, J., McBride, J. M., Koskinen, S., Andersen, J. L., Sipilä, S., Takala, T., & Komi, P. V. (2005). Effects of power training on muscle structure and neuromuscular performance. Scandinavian Journal of Medicine and Science in Sports, 15(1), 58–64.
Markovic, G., & Mikulic, P. (2010). Neuro-Musculoskeletal and Performance Adaptations to Lower-Extremity Plyometric Training. Sports Medicine, 40(10), 859–895.
Mattacola, C., & Dwyer, M. (2002). Rehabilitation of the Ankle after acute Sprain or chronic instability. Journal of Athletic Training, 37(4), 413–429.
Shan, X., Otsuka, S., Yakura, T., Naito, M., Nakano, T., & Kawakami, Y. (2019). Morphological and mechanical properties of the human triceps surae aponeuroses taken from elderly cadavers: Implications for muscle-tendon interactions. PLoS ONE, 14(2).
Turner, A. N., & Jeffreys, I. (2010). The Stretch-Shortening Cycle: Proposed Mechanisms and Methods for Enhancement. Strength and Conditioning Journal, 32(4), 87–99.
Vissing, K., Brink, M., Lønbro, S., Sørensen, H., Overgaard, K., Danborg, K., Mortensen, J., Elstrøm, O., Rosenhøj, N., Ringaard, S., Andersen, J., & Aagaard, A. P. (2008). Muscle adaptations to plyometric vs. resistance training in untrained young men. Journal of Strength and Conditioning Research, 22(6), 1799–1810.
Wilson, J. M., & Flanagan, E. P. (2008). The Role of Elastic Energy in Activities with High Force and Power Requirements: A Brief Review. Journal of Strength and Conditioning Research, 22(5), 1705–1715.
Wu, Y. K., Lien, Y. H., Lin, K. H., Shih, T. T. F., Wang, T. G., & Wang, H. K. (2010). Relationships between three potentiation effects of plyometric training and performance. Scandinavian Journal of Medicine and Science in Sports, 20(1), 80–86.